

Al W. Girotti, PhD
Emeritus Professor
Locations
- Biochemistry
BSB 359
Contact Information
Education
BS, Massachusetts Institute of Technology, 1959
Biography
Dr. Girotti received his Bachelor of Science degree in Biology from Massachusetts Institute of Technology in 1959 and his Doctorate degree in Biochemistry from the University of Massachusetts, Amherst in 1965. He was a Postdoctoral Research Associate at Cornell University Medical College (1965-1968) where he investigated the role of metal ions in ribonuclease activity. Dr. Girotti joined the faculty of the Biochemistry Department at the Medical College of Wisconsin in 1968.
Research Interests
Aerobic cells may experience oxidative stress damage if their enzymatic and non-enzymatic antioxidant defenses are overwhelmed by reactive oxygen species (ROS) generated by various endogenous and exogenous challenges. Unsaturated lipids in cell membranes and lipoproteins are prominent targets of ROS attack, undergoing peroxidative degradation with numerous structurally and functionally disruptive effects. Examples of free radical and non-radical ROS are shown in Scheme 1.
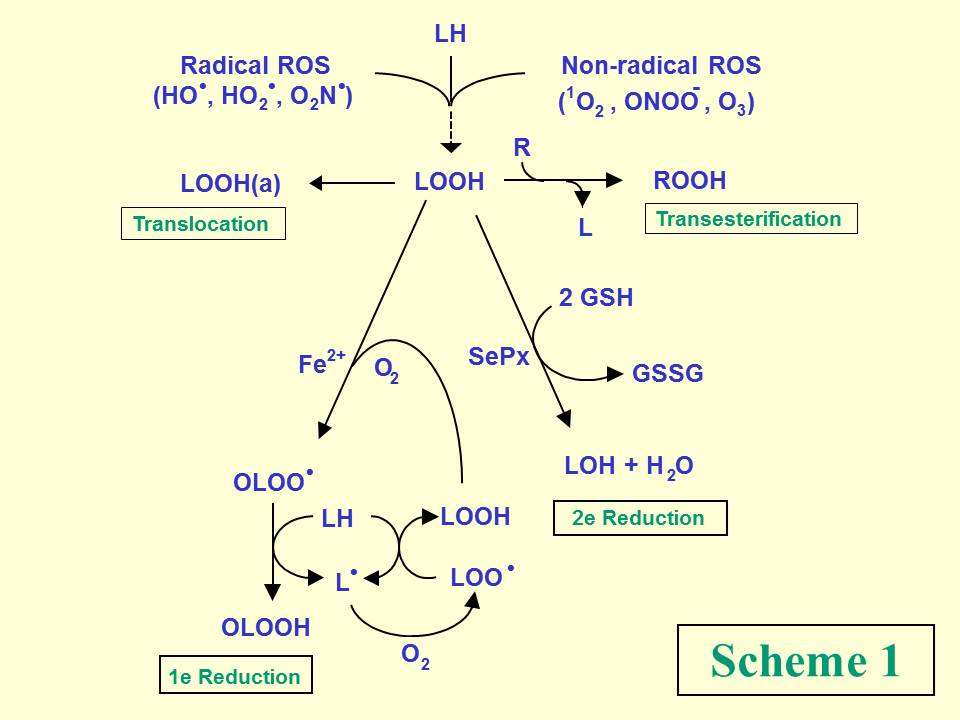
Among the many intermediates/products of lipid peroxidation, hydroperoxide species (LOOHs) are of special interest because of their relatively long lifetimes compared with free radical precursors or products. Under redox-constrained conditions, LOOHs can accumulate steadily with stress duration and may perturb membrane structure/function because of their relatively polar nature. However, in the presence of reductants and catalytic iron, LOOHs can undergo one-electron reduction with formation of oxyl (LO·) and epoxyallylic peroxyl (OLOO·) radicals, which exacerbate membrane damage by triggering chain peroxidation reactions (Scheme 1). Counteracting this is two-electron reductive detoxification catalyzed, for example, by glutathione-dependent selenoperoxidases, GPx4 (also known as PHGPx) being the most prominent isotype. Other LOOH pathways include inter-lipid transesterification and inter-membrane or membrane-lipoprotein translocation.
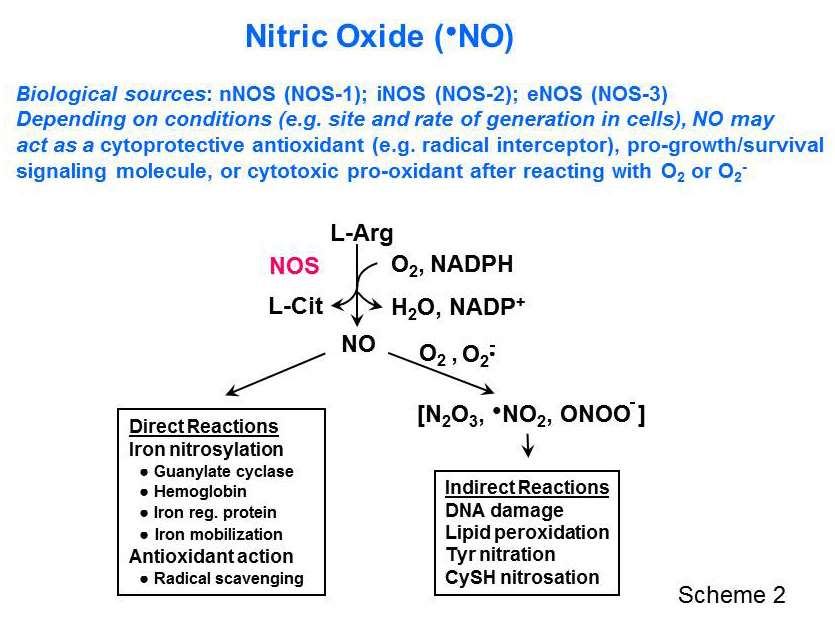
The Girotti group specializes in LOOH formation, turnover, and redox signaling activity, the latter currently attracting widespread biological and biomedical interest. Relatively low LOOH pressure may signal for upregulation of antioxidant proteins and activation of pro-growth transcription factors, whereas high LOOH pressure can signal for growth cessation and programmed cell death (apoptosis). Ongoing projects in the Girotti laboratory include the following: (a) Selenoperoxidase-mediated LOOH metabolism and how this modulates the pathologic as well as therapeutic effects of oxidative stress - as in antitumor photodynamic therapy (PDT), for example; (b) the biological ramifications of spontaneous or transfer protein-facilitated LOOH translocation between membranes or membranes and lipoproteins; pioneering studies of this phenomenon were carried out in the Girotti laboratory; (c) Protective effects of nitric oxide (NO) against cancer cell killing by PDT. Depending on its generation rate and location in cells, nitric oxide synthase (NOS)-derived NO can either be cytotoxic or cytoprotective (Scheme 2). In the latter category, our work has revealed that NO can (i) scavenge lipid-derived free radicals, (ii) induce antioxidant proteins such as heme oxygenase-1 and ferritin, or (iii) engage pro-survival/pro-growth signaling pathways that can compromise the effectiveness of PDT.
Publications
-
(Fil M, Lach M, Bazak J, Sarna M, Girotti AW, Korytowski W.) Photochem Photobiol Sci. 2026 May;25(5):853-864 PMID: 42018124 SCOPUS ID: 2-s2.0-105036346844 04/22/2026
-
(Korbelik M, Girotti AW, Kessel D.) J Mol Med (Berl). 2026 Feb 11;104(1):43 PMID: 41670729 SCOPUS ID: 2-s2.0-105029777982 02/11/2026
-
Preface: Special memorial issue dedicated to Kendric Smith.
(Girotti A, Kessel D, Hasan T, Greer A.) Photochem Photobiol. 2026;102(3):531-532 PMID: 41994922 SCOPUS ID: 2-s2.0-105036108260 04/17/2026
-
(Overchuk M, Choi AM, Rickard BP, Schnoor B, Ehrmann BM, Girotti AW, Zaharoff DA, Huang HC, O'Connell TM, Gomez SM, Rizvi I.) Cell Death Dis. 2025 Dec 10;16(1):899 PMID: 41372113 PMCID: PMC12722221 SCOPUS ID: 2-s2.0-105025463913 12/11/2025
-
(Overchuk M, Choi AM, Wiltshire GAE, Huang HC, Girotti AW, Rizvi I.) J Biomed Opt. 2025 Dec;30(Suppl 3):S34116 PMID: 41503071 PMCID: PMC12770859 SCOPUS ID: 2-s2.0-105026754504 01/08/2026
-
(Girotti AW, Korbelik M.) Photochem Photobiol Sci. 2025 Dec;24(12):1983-1989 PMID: 41251969 SCOPUS ID: 2-s2.0-105022494994 11/18/2025
-
(Krstić L, Vallejo R, Rodriguez-Rojo S, González-García MJ, Arias FJ, Girotti A, Diebold Y.) Int J Pharm. 2025 Jun 10;678:125691 PMID: 40339627 05/09/2025
-
(Korbelik M, Heger M, Girotti AW.) J Lipid Res. 2025 Feb;66(2):100729 PMID: 39675508 PMCID: PMC11911859 SCOPUS ID: 2-s2.0-105000501101 12/16/2024
-
Tumor Lipid Signaling Involved in Hyperoxidative Stress Response: Insights for Therapeutic Advances.
(Korbelik M, Girotti AW.) J Cell Signal. 2025;6(2):39-47 PMID: 40458196 PMCID: PMC12129048 06/03/2025
-
(Lapoot L, Jabeen S, O'Connor RM, Korytowski W, Girotti A, Greer A.) J Org Chem. 2024 Sep 20;89(18):12873-12885 PMID: 39231123 PMCID: PMC11421024 SCOPUS ID: 2-s2.0-85203176696 09/04/2024
-
(Girotti AW, Korytowski W.) Int J Mol Sci. 2024 May 23;25(11) PMID: 38891885 PMCID: PMC11171770 SCOPUS ID: 2-s2.0-85195836378 06/19/2024
-
(Pabisz P, Bazak J, Sabat M, Girotti AW, Korytowski W.) Cell Biochem Biophys. 2024 Mar;82(1):213-222 PMID: 37995086 PMCID: PMC10866752 11/23/2023